

Which Part Of The Sarcomere Stays Almost The Same Size During Contraction
Introduction and Background
Muscle contraction has fascinated lay people and scientists for centuries. Still, a good understanding of how muscle contraction occurs seemed but possible once microscopy techniques had evolved to a level where basic structural features, such equally the regular cross striation patterns of fibers, could exist observed in the belatedly 19th century. In the early 20th century, a stimulated musculus was just considered a new rubberband body (Gasser and Hill, 1924). Shortening and work product took identify with a fixed amount of energy that was stored in this body and evolved elastically through stimulation. Yet, this notion was proven false when Wallace Fenn demonstrated that muscle produced an increasing amount of total energy when increasing its mechanical piece of work output; an observation that was in contradiction with Hill'southward elastic body theory (Fenn, 1923, 1924). Specifically, Fenn, who worked in the laboratory of Hill and measured estrus and work production in frog muscles, found that a muscle allowed to shorten liberated more than energy than a muscle held isometrically or a musculus that was stretched. This has become known every bit the Fenn result in muscle physiology.
Prior to the 1950s, muscle contraction and force production were thought to be caused by the folding of long protein chains visible in the heart of the sarcomere. This shortening had been idea to exist caused by lactic acid, but this theory was refuted past experiments demonstrating that contractions could be obtained in the absence of lactic acid in muscles poisoned with iodoacetic acid (Lundsgaard, 1930). The role of lactic acrid was then replaced briefly with phosphorylcreatine breakdown, until it was discovered that this reaction merely served to re-phosphorylate ADP into ATP. Thereafter, filament shortening became associated with the hydrolysis of ATP into ADP and inorganic phosphate.
In the early 1950s, careful analysis of A-ring dimensions revealed that myosin filaments were non substantially shortening under a diversity of contractile weather, and thus, could not account for muscle wrinkle, force production and the big length changes that muscle tissue tin undergo (Huxley, 1953). In two seminal papers, arrived at independently, Hugh Huxley and Andrew Huxley proposed that musculus contraction occurred not by shortening of the myosin filaments, only by the relative sliding of two sets of filaments, actin, and myosin (Huxley and Hanson, 1954; Huxley and Niedergerke, 1954). In 1957, Andrew Huxley proposed how this relative sliding might occur, and provided a mathematical framework for what is now known every bit the cross-span theory of muscle wrinkle (Huxley, 1957). This paper, which has been cited over 3000 times (Google Scholar, June 2014), outlines in broad strokes how nosotros recall about muscle contraction today. The success of this paper is insofar surprising as Huxley never intended to publish information technology, and thought of the mathematical formulation of the cross-bridge theory merely as a preliminary idea (Huxley, personal advice, Baronial 1999). He sent the paper to his friend and editor of Biophysics and biophysical Chemistry, who, to Huxley's surprise, suggested publishing information technology.
Unaccounted Observations
The initial two land model of the cantankerous-bridge theory published in 1957 underwent several reformulations, although the basic premise remained unchanged. Briefly, in the cross-bridge model, contraction and force production is accomplished by extensions (cantankerous-bridges) from the thick (myosin) filaments that interact cyclically with the sparse (actin) filaments and exert forcefulness between these two sets of filaments to produce shortening. Each wheel of zipper and detachment of a cross-bridge is associated with the hydrolysis of one molecule of ATP. Therefore, the regulation of force is governed exclusively by the contractile proteins actin and myosin, while structural proteins provide passive forces upon muscle elongation that are determined exclusively by their viscoelastic backdrop.
In 1969, the cross-bridge theory was amended with the idea that cross-bridges produce force and shortening through rotation requiring multiple attached states (Huxley, 1969; Huxley and Simmons, 1971), and in 1993, a detailed clarification of the diminutive structure of cross-bridges and the corresponding actin zipper sites revealed a cross-bridge stroke that included rotation of the cross-span around a fixed element of the cantankerous-bridge head that attached uniquely to the actin zipper site (Figure ane) (Rayment et al., 1993).
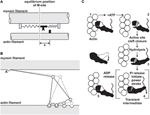
Figure i. Development of the cross-bridge model of muscle contraction. (A) Original ii-country model proposed by Huxley (1957). (B) Multi-state cantankerous-bridge model with rotating head as proposed initially by Huxley (1969) and mathematically described past Huxley and Simmons (1971). (C) Multi-state model based on the diminutive structure of cross-bridges and actin attachment sites as proposed by Rayment et al. (1993).
In his original clarification of the cross-bridge theory, Huxley (1957) was able to predict forces for concentric contractions accurately. Specifically, Huxley derived a set of rate constants for the attachment/detachment kinetics of cross-bridges that accurately predicted "the best available information at the time," the concentric force-velocity human relationship of striated muscles (Hill, 1938). Furthermore, the cross-bridge theory besides explained beautifully the isometric forcefulness as a role of cobweb and sarcomere lengths (Gordon et al., 1966). However, the forces and energetics for eccentric contractions (actively stretched muscles), were not predicted accurately: they were much too big (Huxley, 1957). Also, the well-known results on residual force enhancement following agile stretching of muscles (Abbott and Aubert, 1952; Edman et al., 1982) could not be predicted conceptually (Walcott and Herzog, 2008) or numerically (Herzog et al., 2012a,b; Herzog, 2014).
Residual forcefulness enhancement is an best-selling belongings of skeletal muscle (Edman et al., 1982). Information technology describes the increase in steady-state isometric strength following an active musculus stretch, compared to the corresponding purely isometric force at the same length and aforementioned activation (Figure 2). Remainder force enhancement has been observed in whole muscle preparations, activated voluntarily (Oskouei and Herzog, 2005) or through electrical stimulation (Lee and Herzog, 2002), in single intact and skinned fibers (Edman et al., 1978, 1982; Sugi and Tsuchiya, 1988; Rassier et al., 2003c; Peterson et al., 2004; Lee and Herzog, 2008; Joumaa and Herzog, 2013), in myofibrils (Rassier et al., 2003a; Joumaa et al., 2008) and in single, mechanically isolated sarcomeres (Leonard et al., 2010) and half sarcomeres (Joumaa et al., 2008). Rest force enhancement cannot exist predicted using the cross-bridge theory (Walcott and Herzog, 2008), because the rate constants of cantankerous-bridge zipper/detachment do not depend on time but only on the relative location of the cross-bridge's equilibrium position relative to its nearest attachment site on actin (Huxley, 1957). Therefore, an explanation for balance strength enhancement needed to be found that would not undermine the cross-bridge theory. Thus, for more than half a century, residual strength enhancement was explained conceptually, albeit not numerically, with the idea of instability of sarcomeres on the descending limb of the force-length human relationship (Hill, 1953) and the associated evolution of large sarcomere lengths non-uniformities (Morgan, 1990, 1994).

Figure 2. Residuum strength enhancement in skeletal muscles. (A) Residual force enhancement (FE) and passive strength enhancement (PFE) in a whole muscle preparation (true cat soleus at 37°C). (B) Passive forcefulness enhancement (PFE) in a single myofibril training (rabbit psoas at 21°C). (C) Strength enhancement (Fe) and force above the isometric plateau (O-Iron) in a single sarcomere preparation (rabbit psoas at 21°C). The gray line in (A) represents the isometric reference contraction while the black lines represent active stretch contractions followed past an isometric contraction. The gray line in (B) represents passive strength during myofibril stretching and the black line the corresponding active strength; deactivation occurred at about 55 s. The greyness line in (C) represents the isometric reference force while the black line represents the experimentally enhanced force post-obit an active stretch.
Caption of Remainder Force Enhancement (Using Sarcomere Length Non-Uniformity)
According to the sarcomere length non-uniformity theory, sarcomeres on the descending limb of the force-length relationship are unstable (Hill, 1953; Allinger et al., 1996; Zahalak, 1997). This instability is thought to exist caused by a "weakening" behavior of sarcomeres (negative stiffness). Therefore, for a perturbation, such as active stretching of muscle on the descending limb of the force-length human relationship, sarcomeres were thought to exist destabilized, causing a quick, uncontrolled over-stretching (popping) of some sarcomeres at the expense of others that simply stretch slightly, non at all, or might even shorten by a small corporeality. The popped sarcomeres were thought to attain strength equilibrium with the curt (active) sarcomeres through passive forces that become high at long lengths. Forcefulness enhancement was so explained with the idea that isometric contractions on the descending limb do not produce a sufficient perturbation to sarcomeres, thus sarcomeres remain relatively uniform and thus produce a strength in accordance with actin-myosin filament overlap (Gordon et al., 1966). In dissimilarity, a muscle that is stretched actively was idea to produce perturbations that event in instabilities and large sarcomere length non-uniformities that requite ascension to 2 distinct sets of sarcomere lengths. The steady-state force following active stretch was then idea to be greater than the purely isometric forcefulness because the agile sarcomeres are shorter following agile stretch compared to the purely isometric contraction (and thus can produce more strength), and the passive sarcomeres are pulled to such lengths that their passive forces friction match the forces of the short, active sarcomeres (Figure iii).
Effigy 3. Force enhancement (FE) based on the sarcomere length non-uniformity theory. Isometric contractions of muscles on the descending limb of the force-length relationship are idea to occur with sarcomeres of compatible lengths (blackness circle and grey foursquare). However, if a muscle is stretched from a short length (blackness circle) to a long length (grey foursquare), some sarcomeres are thought to become overstretched (popped—right black diamond) while others are idea to be stretched just minimally (left black diamond), thus producing an average sarcomere length the same as that of the long isometric wrinkle (gray square), but producing more force because of the more favorable position of the curt sarcomeres (left black diamond) and the passive forces of the overstretched sarcomeres (right black diamond).
In the following, we would like to identify predictions that are direct outcomes of the mathematical conception of the sarcomere length not-uniformity theory. We will discuss these theoretical predictions in view of existing experimental prove, and then will attempt to draw conclusions about the ceremoniousness of this theory.
Predictions Based on the Sarcomere Length Non-Uniformity Theory
The theoretical models of the sarcomere length non-uniformity theory provide uniquely testable predictions (Zahalak, 1997; Campbell, 2009). Some of the primary predictions have been identified and discussed by proponents and opponents of this theory (Morgan et al., 2000; Herzog and Leonard, 2006, 2013; Herzog et al., 2006; Morgan and Proske, 2006; Edman, 2012). However, other equally obvious predictions, have received less attention, peradventure because of the difficulties in testing them.
The primary predictions of the sarcomere length not-uniformity theory that have been discussed in previous works might be summarized equally follows:
1. Sarcomeres are unstable on the descending limb of the force-length relationship following active stretching (Morgan, 1990, 1994).
2. Instability of sarcomere lengths, and thus the evolution of sarcomere length non-uniformities (and associated strength enhancement) can just occur on the unstable descending limb but not the stable ascending limb of the force-length relationship (Allinger et al., 1996; Zahalak, 1997).
3. Forces in the force enhanced state cannot exceed the purely isometric forces at the plateau of the force-length relationship (Edman et al., 1982; Rassier et al., 2003c).
iv. Force enhancement cannot occur in a single sarcomere preparation (Leonard et al., 2010).
Secondary predictions that follow directly from the mathematical framework and conceptual thinking of the sarcomere length non-uniformity theory that have received less, or no attention, may be summarized as follows:
5. Isometric contractions on the descending limb of the strength-length relationship are associated with sarcomeres of essentially uniform length (Morgan, 1990, 1994).
six. Sarcomere length non-uniformities increase when a muscle is actively stretched compared to the corresponding purely isometric contractions (Morgan, 1990, 1994; Allinger et al., 1996).
7. Sarcomere lengths following agile musculus stretching will have two distinct values (Allinger et al., 1996; Walcott and Herzog, 2008).
The primary predictions (one–4) of the sarcomere length non-uniformity theory have been discussed extensively (Herzog et al., 2006, 2012a,b; Edman, 2012; Herzog, 2014) just for completeness are summarized here briefly.
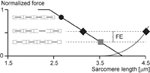
1. Instability of sarcomeres on the descending limb of the strength-length relationship: More than 30 years ago, when first reading about sarcomere length instability on the descending limb of the force-length relationship, I asked myself the question: why would nature evolve a universal motor for contraction that was unstable and would tear itself apart, over more than than one-half of its potential working range? After developing a setup for single myofibril testing, information technology was the first question I wanted to be answered. Stretching of serially arranged sarcomeres onto the descending limb of the force-length relationship did not produce sarcomere length instabilities (Rassier et al., 2003b); i.e., at that place was no quick, uncontrolled popping of the "weakest" sarcomeres, as predicted by the theory (Morgan, 1990, 1994). Rather, sarcomeres were perfectly stable at vastly differing lengths on the descending limb of the force-length relationship (Figure 4A), an observation that still needs satisfactory caption. Oftentimes, sarcomeres that were shorter compared to other sarcomeres prior to active stretching, were longer after stretching (Figure 4B), a finding that is incompatible with the sarcomere length non-uniformity and cross-bridge theories.
two. Force enhancement on the ascending limb of the strength-length relationship: The ascending limb of the forcefulness-length human relationship has a positive slope, and thus strengthening character which produces inherent sarcomere length stability (Epstein and Herzog, 1998). Therefore, co-ordinate to the sarcomere length non-uniformity theory, at that place should be no force enhancement on the ascending limb of the force-length relationship. However, in the very first systematic analysis of force enhancement, Abbott and Aubert (1952) reported force enhancement on the ascending limb of isolated muscle preparations. This initial finding was supported by farther observations on whole muscles (Morgan et al., 2000), and unmarried fibers (Peterson et al., 2004). Notwithstanding, force enhancement on the ascending limb tends to be small compared to the descending limb, and thus, although consistently observed, may not always exist acknowledged (Morgan et al., 2000; Edman, 2012).
3. Enhanced force above the isometric plateau forces: In the sarcomere length non-uniformity theory, the steady-state isometric forces, independent of the history of contraction, cannot exceed the isometric forces obtained at the plateau of the forcefulness-length relationship. Since, the brusk sarcomeres in this theory are the active force producers, and since they must exist within a region of actin-myosin filament overlap, the maximal force they tin can produce is that obtained at optimal sarcomere length represented by the plateau of the force-length relationship (Gordon et al., 1966). Yet, forces in the enhanced state exceeding those of purely isometric contractions at optimal lengths take been observed as early on as 1978 (Edman et al., 1978), although these authors later revised their results. Nonetheless, enhanced forces conspicuously exceeding the isometric plateau forces were found in a series of subsequent studies on whole muscles (Schachar et al., 2004), single fibers (Rassier et al., 2003c; Lee and Herzog, 2008), and most importantly, in unmarried myofibrils and single, mechanically isolated sarcomeres (Leonard et al., 2010) (Figure 2). In conclusion, isometric steady-land forces following active muscle stretch can exceed the purely isometric forces at optimal length past a substantial amount.
iv. Forcefulness enhancement in a single sarcomere: Obviously, if strength enhancement requires the evolution of sarcomere length not-uniformities, every bit indicated in Effigy 3, then force enhancement should never occur in a single sarcomere. The classic work by Tim Leonard was the offset published research on force enhancement in a mechanically isolated single sarcomere preparation (Leonard et al., 2010). In that report, x unmarried sarcomeres were isolated and stretched from optimal length (2.4 μm) to a length of 3.four μm, and compared to the steady-land forces obtained for purely isometric contractions. Strength enhancement was 190% on boilerplate and the enhanced forces exceeded the purely isometric forces at optimal length by an average of 37%. It has been suggested that these results might take been obtained due to the evolution of half-sarcomere length non-uniformities in this unmarried sarcomere grooming. However, if and then, the question remains how a single one-half-sarcomere can produce forces in excess of its isometric forcefulness at optimal length. Needless to say that one must be careful of results from a single written report that have non been repeated in other laboratories, only for lack of evidence to the contrary, we take that strength enhancement is a sarcomeric property, and can occur in the absence of multiple sarcomeres of vastly different lengths.
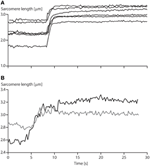
Figure 4. Sarcomere length stability on the descending limb of the force-length relationship. (A) Sarcomere lengths as a function of time for a six sarcomere myofibril stretched on the descending limb of the force-length relationship. Note that none of the sarcomeres is chop-chop stretched beyond actin-myosin filament overlap (popped approximately 4.0 μm), but that they stay relatively constant in length for a xx s flow post-obit agile stretching. (B) Sarcomere lengths of two specific sarcomeres from a single myofibril. Note, when the myofibril is stretched, the initially short sarcomere becomes the long sarcomere and vice versa.
The secondary predictions (5–7) of the sarcomere lengths non-uniformity theory have received much less, or no attention in the past, just seem equally relevant and will be discussed in the following.
5. Compatible sarcomere lengths for isometric contractions: For the sarcomere length non-uniformity theory to work, specifically for it to account for the residual forcefulness enhancement property of skeletal muscle, it is necessary that sarcomeres are essentially uniform for isometric contractions. Agile stretching is then thought to be the stimulus that produces sarcomere length instability and associated length not-uniformities. At that place take been extensive reports that sarcomere lengths in muscle fibers are highly not-uniform (Huxley and Peachey, 1961), thus requiring specialized approaches when studying sarcomere force-length properties (Gordon et al., 1966). Sarcomere length non-uniformities take been primarily observed as boilerplate sarcomere length variations across unmarried fibers, but more than recently have been demonstrated for single sarcomeres in whole muscles (Llewellyn et al., 2008), and in single myofibrils (Figure 4). When quantifying sarcomere lengths in passive and agile human being muscles, Llewellyn et al. (2008) noticed variations in sarcomere lengths of xx% in a radius as minor every bit 25 μm, while nosotros institute peak sarcomere length not-uniformities for purely isometric contractions of 37% in isolated myofibril preparations. Sarcomere lengths variations have been shown to range from 1.7 to 3.five μm in frog semitendinosus fibers at rest (Huxley and Peachey, 1961). Taken together, these results suggest that sarcomere lengths non-uniformities are a natural occurrence of resting and activated muscle preparations at all structural levels. Thus, observing sarcomere length non-uniformities after stretch or shortening contractions should non imply that the dynamics of muscle wrinkle produced these non-uniformities, nor should these not-uniformities be thought to be the cause of specific mechanical properties of musculus without careful analyses.
6. Increment in sarcomere length non-uniformity with active stretching of musculus: One of the tenets of the sarcomere length non-uniformity theory in explaining force enhancement is the idea that sarcomere lengths go non-uniform during agile stretching on the descending limb of the strength length relationship, while they remain uniform with passive stretching and subsequent isometric contraction. If sarcomere lengths are indeed more than non-compatible in the force enhanced compared to the isometric reference state has never been tested systematically. Initial work on this topic was done by stretching active single fibers or whole muscle preparations, and fixing them quickly following the stretch. These "stretched" preparations were then compared histologically to corresponding preparations that were activated isometrically or were allowed to actively shorten (Julian and Morgan, 1979; Morgan et al., 1982). These experiments typically showed an increased number of overstretched (popped) sarcomeres in the actively stretched muscles compared to those not undergoing active lengthening. These experiments have the advantage that they are performed in intact preparations, but have the disadvantage that overstretching of the sarcomeres cannot be accounted for past instability, rather than, for example, pre-existing structural impairment of the musculus, and that only a tiny fraction of the whole muscle was analyzed for overstretched sarcomeres, thus making generalization hard.
Reports of stability or sarcomere lengths non-uniformities later active stretching propose that, if anything at all, sarcomeres are more stable (Edman et al., 1982), and sarcomere lengths are more than uniform post-obit active muscle stretching compared to the corresponding purely isometric reference contractions (Joumaa et al., 2008). Ongoing work in our lab has focused on measuring sarcomere length non-uniformities in isolated myofibril preparations in the force enhanced and normal isometric reference states. Preliminary results propose that there is no systematic increase in sarcomere length non-uniformity, as quantified past the range and variance of sarcomere lengths post-obit eccentric contractions on the descending limb of the force-length relationship compared to the corresponding purely isometric reference contractions.
7. Two singled-out sarcomere lengths following active muscle stretching: In a muscle or cobweb grooming, sarcomeres are not only connected serially but also in parallel. Therefore, forcefulness transmission by a sarcomere is not just adamant by the force information technology exerts, only also by the forces acting on it in serial and in parallel. However, in an idealized preparation, as typically modeled theoretically, sarcomeres are causeless to be perfectly in series. Such an idealization is accomplished experimentally when using single myofibril preparations, which makes this preparation uniquely attractive to study sarcomeric properties. According to the sarcomere length not-uniformity theory, sarcomeres in series (in a myofibril) should be separated into two distinct groups with 2 singled-out lengths when actively stretched. I group representing the sarcomeres that were not or only slightly stretched, the other representing the overstretched (popped) sarcomeres. Quantification of sarcomere lengths from a variety of published studies suggest that sarcomere lengths practice non fall into two distinct lengths categories, but rather, are distributed over a range of lengths (eastward.g., Effigy 4). Conscientious analysis of the sarcomere lengths of twelve myofibrils did not reveal a single one of them having ii distinct sarcomere lengths in the forcefulness enhanced state thus defying this specific prediction.
Further Considerations
Force-Length Relationship
1 of the most puzzling results of muscle physiology is the different shapes of the descending limbs of the force-length relationships for and so-called "stock-still-cease" and "segment-clamped" conditions (Effigy v). In fixed-cease experiments, the two ends of a fiber are stock-still to a motor and a force transducer, respectively, and measurements of isometric force are made as a function of fiber lengths (Pollack, 1990). In segment-clamped experiments, a pocket-sized mid-section of a cobweb with relatively uniform sarcomeres is identified and marked, and its length is kept abiding using length feedback by carefully adjusting the entire fiber length. Isometric forces are and then expressed as a function of the sarcomere lengths in the fixed section of the fiber. For the fixed-end contractions, sarcomeres are immune to have on their "normal" not-uniform length distribution (Pollack, 1990), while in the segment-clamped arroyo, sarcomere lengths are kept constant for the clamped segment (Gordon et al., 1966). Isometric forces on the descending limb of the force-length relationship in the stock-still-end contractions have been reported to be substantially greater than those obtained in the corresponding segment-clamped experiments. Therefore, information technology appears that allowing sarcomeres to attain their natural non-uniform length distribution during purely isometric wrinkle enhances force, while attempting to go along sarcomere lengths artificially uniform results in a substantial loss of isometric force. Nosotros may conclude from these results that even during isometric contractions, sarcomere length non-uniformities develop naturally, they practise non depend on agile stretching on the descending limb of the force-length relationship, and they, in some unknown fashion, contribute to increased isometric force.
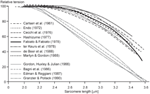
Effigy v. Descending limb of the sarcomere force-length relationship. Relative tension every bit a function of sarcomere lengths obtained from stock-still end contractions (blackness lines) and from segment-clamped experiments (gray lines). Note that when the natural development of sarcomere length non-uniformities is prevented by segment clamping, the isometric forces are severely decreased (Pollack, 1990).
Theoretical Force-Length Relationship of Single Sarcomeres
When it was first demonstrated that force enhancement could exist observed in a unmarried sarcomere preparation (Leonard et al., 2010), critics were quick to point out the possibility that force enhancement could have been caused by the development of one-half-sarcomere non-uniformities. Although this argument sounds appealing on the surface, its acceptance leads quickly to a variety of unlikely consequences. Only i of these shall be discussed here.
Imagine nosotros have a single sarcomere just at the border of the plateau leading to the descending limb of the force-length relationship (Effigy half dozen) and we now stretch this sarcomere. For the sake of argument, let'south also assume that at this initial length, there is no passive force (or if at that place was, we could subtract it from the initial considerations without affecting the following statement). Imagine now that we stretch this sarcomere, so its total length is ¼down, then ½down, and then at the full length downwards on the descending limb of the strength-length relationship. If we now presume that one one-half of the sarcomere remains at its initial short length, then the other half would have to elongate twice the stretch magnitude of the whole sarcomere (Effigy six), and, for force equilibrium'due south sake, its strength has to be equal to that of the one-half sarcomere that remained at its initial length. For this to occur, the passive force would have to start at the beginning (left manus side) of the descending limb of the force-length relationship, it would have to be a direct line, and its slope would have to be the same as that of the descending limb, except with opposite sign (positive instead of negative). Then, when the half sarcomere loses actin-myosin filament overlap, the passive forcefulness would accept to remain constant. Needless to say, such a passive force has never been observed, and even if it had, it withal could not explain the results by Leonard et al. (2010), where forces in the enhanced state clearly exceeded the isometric plateau forces in a unmarried sarcomere preparation. This case illustrates how ane has to be conscientious when making a proposal, or stating a critique, without thinking through the associated implications.
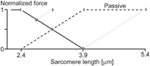
Figure half dozen. Theoretical strength-length relationship of a unmarried sarcomere. If we presume that force enhancement in a single sarcomere, as observed in the literature (Leonard et al., 2010), is acquired by one-half-sarcomere length not-uniformities, and that active strength is proportional to actin-myosin filament overlap, so the passive force would have to look as indicated in the figure (thick black dashed line). Such a passive force has never been observed and seems to contradict anything known about passive forces in skeletal muscles. Therefore, the notion that substantial force enhancement in a unmarried sarcomere tin be explained with the development of half-sarcomere length not-uniformities seems far-fetched, not to say impossible. Total circles bespeak an entire sarcomere on the descending limb of the force-length relationship, as expected during an isometric wrinkle with equal half sarcomeres. Half-circles indicate the corresponding half-sarcomeres expected later an active stretch in the force enhanced state of a single sarcomere. Black line = descending limb of the active force-length relationship. Dashed black line = passive force required for the overextended half-sarcomere to friction match the forcefulness of the other, agile, half sarcomere.
A-Band Shifts and Forcefulness Enhancement
In a recent written report, A-band shifts, which indicate the degree of half-sarcomere length non-uniformities, were argued to "significantly increase the level of force enhancement (Rassier, 2012)." The argument was based on observed correlations of "stretch-induced" A-ring shifts with residual force enhancement in single myofibril preparations (Figure 7). Here, a maximal A-band shift of approximately 72 nm was associated with an enhanced strength of about 55%. However, a 72 nm shift would explain a 10% increase in strength at best, thus well-nigh of the enhanced strength remains unexplained. In a like effigy on single sarcomere experiments, with an implied spatial resolution of less than 0.v nm, force enhancement for a 12 nm A-band shift was approximately xx%. A 12 nm shift explains a one.five% increase in force leaving 92.five% of the observed force enhancement unexplained.
Effigy seven. Force enhancement as a function of half-sarcomere length non-uniformities. (A) Force enhancement as a function of A-band displacements (equivalent to half sarcomere length non-uniformities) in single sarcomere preparations, and (B) in single myofibril preparations with multiple serial sarcomeres. Note that the small-scale A-band shifts in the single sarcomere and the single myofibril only explain approximately 7 and 18% of the full force enhancement observed, assuming that the isometric reference contractions take perfectly identical half- and sarcomere length. Yet, since the isometric reference contractions accept similar A-band shifts every bit the experimental agile stretch contractions, none of the enhanced forces seems explained by stretch-induced half-sarcomere length non-uniformities from Rassier (2012) with permission.
Moreover, experiments with no A-band shifts were found to have top strength enhancements of upwardly to 16% (Figure 7). Finally, and probably nigh telling, half-sarcomere length non-uniformities for the isometric reference contractions, although non systematically evaluated in this study, were greater (their Figure 5C) or equal (their Figure 6C) to the not-uniformities of the actively stretched experimental contractions, thus it is hard to support the authors' claim that the force enhancement was associated with "stretch-induced" non-uniformities of one-half sarcomere lengths. Albeit not systematically evaluated, in studies where half-sarcomere length non-uniformities were compared for isometric reference and experimental stretch contractions, one-half-sarcomere lengths tended to be more compatible afterwards stretch compared to reference contractions (Joumaa et al., 2007). Combined, these results provide fiddling support that stretch-induced one-half-sarcomere non-uniformities contribute to the residual force enhancement in skeletal muscles.
Conclusions and Future Directions
Ever since the emergence of the cross-bridge theory, properties of actively stretched muscles could not be predicted properly (Huxley, 1957). Forces and energy consumption were much as well big compared to experimental results (Huxley, 1957), and residual force enhancement could not be predicted conceptually, as steady-state forces in the cross-bridge theory are independent of the history of contraction (Huxley, 1957; Huxley and Simmons, 1971; Walcott and Herzog, 2008). This shortcoming of the cross-bridge theory had been addressed past assuming that musculus segments (Hill, 1953) and sarcomeres (Morgan, 1990, 1994) were unstable on the descending limb of the strength-length relationship, and small-scale perturbations would crusade cracking not-uniformities in sarcomere lengths. Active lengthening of muscles (eccentric contraction) was thought to be such a perturbation. It is interesting to notation that the sarcomere length instability and associated stretch-induced development of sarcomere length non-uniformity theory has survived for such a long time, and in many circles is notwithstanding unquestionably accepted, despite lack of straight evidence, and despite experimental results from the very first that were not in agreement with the predictions of the theory. For example, Abbott and Aubert (1952) had strong evidence of force enhancement on the ascending and plateau regions of the force-length relationship more than than half a century ago, predating the formulation of the cross-bridge theory itself.
The refinement of mechanical experiments on unmarried myofibrils (Bartoo et al., 1993; Rassier et al., 2003a; Joumaa et al., 2008; Leonard et al., 2010; Leonard and Herzog, 2010) and mechanically isolated sarcomeres (Leonard et al., 2010) has allowed for direct testing of many of the predictions of the sarcomere length non-uniformity theory. Among these rejected predictions (see word above), the ones most dissentious to the non-uniformity thinking were the following:
1. The evidence that sarcomeres of vastly different length could reside side by side on the descending limb of the strength-length human relationship without observable length changes over periods of 30 s, thereby demonstrating stability;
2. That substantial force enhancement could occur in a unmarried, mechanically isolated sarcomere;
iii. And that the enhanced forces in single sarcomeres could exceed the purely isometric reference forces obtained at optimal sarcomere length by a substantial corporeality.
There is no doubtfulness that sarcomeres in the forcefulness enhanced state are not-compatible. Still, sarcomeres in muscles (Llewellyn et al., 2008), fibers (Huxley and Peachey, 1961) and myofibrils (Rassier et al., 2003a; Joumaa et al., 2008; Leonard and Herzog, 2010; Leonard et al., 2010) are also not-uniform for purely isometric contractions. Whether, or not these sarcomere length-non-uniformities increment with stretching has not been systematically elucidated, but pilot results, and isolated findings from unrelated studies, suggest that, if anything at all, sarcomeres following active muscle stretching are more stable (Edman et al., 1982) and accept equal or less sarcomere length-non-uniformity compared to the corresponding isometric reference contractions (Joumaa et al., 2008; Rassier, 2012).
From these results, and testify in the literature, we propose that sarcomere length non-uniformities are a normal acquaintance of muscle contraction on all structural levels. They are not an occurrence sectional to agile muscle stretching (eccentric contractions) on the descending limb of the forcefulness-length relationship, and they are not a primary cause for the enhanced forces observed in skeletal muscles after active stretching.
If not the development of stretch-induced sarcomere length non-uniformities, how else can we explicate residuum force enhancement? A lilliputian over a decade ago, nosotros discovered the existence of a passive component of rest force enhancement (Herzog and Leonard, 2002). Testify accumulated over the by decade strongly suggests that the structural protein titin causes this passive strength enhancement (Herzog et al., 2006). The idea was that titin, which acts equally a molecular jump in the I-ring region of sarcomeres, alters its jump stiffness when a muscle is activated. In principle, titin's stiffness can be changed in two ways: (i) by changing its material backdrop, or (ii) past changing its free spring length. Recent findings support the thought that both these mechanisms are at work in actively stretched skeletal muscles (Figure viii).
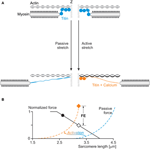
Figure eight. (A) Titin-induced force enhancement: a one-half-sarcomere is stretched passively (left) and actively (right). During passive stretching, titin elongates according to its normal spring properties. During agile stretching, calcium binds to titin and titin's proximal region binds to actin: both of these events increase titin'southward stiffness and thus its force when actively stretched compared to when passively stretched. (B) The effects of activation on titin'due south force (passive force) are illustrated schematically in the strength-length graph with a shift of the passive force to the left of the sarcomere length calibration, and an increase in stiffness at a given sarcomere length.
Titin is known to increment its stiffness upon muscle activation by binding calcium to specialized sites (Labeit et al., 2003; Joumaa et al., 2007). Labeit et al. (2003) identified the glutamate rich-region of the so-called PEVK segment of titin as a binding site for calcium, and Duvall et al. (2012) showed that calcium binds to specific Immunoglobulin (Ig) domains of titin and by doing so makes titin stiffer. There might exist other calcium bounden sites on titin that have however to be identified.
Titin is also thought to change its gratis jump length by binding its proximal region to actin upon activation and muscle force production. Leonard and Herzog (2010) demonstrated that actively stretched sarcomeres and myofibrils produced virtually 3–4 times greater forces than passively stretched myofibrils in regions where actin-myosin filament overlap had been lost, and thus agile forces were zero. Similarly, early findings on labeled titin segments in myofibrils indicate that during passive stretching, all I-band segments of titin elongate, as expected, while during agile stretching, some proximal segments in the I-band region do not elongate, suggesting that they may be bound to a rigid backbone, for example the actin filament. In mdm knockout mice, where role of the N2A region of titin, thought to be critical for attachment of titin to actin, is eliminated, the difference between passive forces in actively and passively stretched myofibrils is small (about xv%) and can readily be explained with calcium binding to the titin segments identified above.
These results point to titin as a force regulating poly peptide in muscle contraction; specifically in eccentric contractions and in the force enhanced state. Corresponding conceptual models of such a three filament strength regulating sarcomere (Figure 8) accept been discussed elsewhere in detail (Herzog et al., 2012a,b; Herzog, 2014) and a corresponding mathematical model has been developed and recently published (Schappacher-Tilp et al., 2015).
Independent of the ultimate caption for residual force enhancement in skeletal muscles, the proposed substitution of the two filament (actin and myosin) with a three filament (actin, myosin, and titin) sarcomere model for force production has a variety of advantages over the sarcomere length non-uniformity theory, not the least of which is that it tin can explicate all isometric and concentric force properties of the traditional cross-bridge model, but can as well predict the increased forcefulness during eccentric contractions, the efficiency of eccentric contractions and the agile and passive strength enhancement following agile muscle stretching.
But not only that, the three filament model of muscle force product has some intuitively appealing properties, including:
i. The passive structures of muscles are soft and compliant when passively stretched merely become hard and potent during active stretching, thus providing boosted force at negligible energetic cost.
ii. Titin forces increase when actin-myosin forces decrease, thereby providing a machinery preventing damage in muscles stretched actively to long lengths.
iii. Titin provides stability for sarcomeres on the descending limb of the strength-length relationship and for myosin filaments in the center of the sarcomere. When titin is eliminated, all passive, and agile strength transmission across sarcomeres is lost (Leonard and Herzog, 2010).
Disharmonize of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or fiscal relationships that could be construed as a potential disharmonize of interest.
Acknowledgments
The Canadian Institutes of Health Research (CIHR), the Natural Sciences and Applied science Inquiry Council of Canada (NSERC), the Killam Foundation, and the Canada Research Chair Program for Molecular and Cellular Biomechanics.
References
Abbott, B. C., and Aubert, X. M. (1952). The strength exerted by agile striated muscle during and afterwards alter of length. J. Physiol. 117, 77–86.
PubMed Abstract | Google Scholar
Allinger, T. L., Epstein, M., and Herzog, W. (1996). Stability of musculus fibers on the descending limb of the force- length relation. A theoretical consideration. J. Biomech. 29, 627–633. doi: x.1016/0021-9290(95)00087-ix
PubMed Abstract | CrossRef Full Text | Google Scholar
Bartoo, G. Fifty., Popov, V. I., Fearn, L. A., and Pollack, G. H. (1993). Active tension generation in isolated skeletal myofibrils. J. Musculus Res. Cell Motil. fourteen, 498–510. doi: 10.1007/BF00297212
PubMed Abstract | CrossRef Full Text | Google Scholar
Campbell, K. S. (2009). Interactions between connected half-sarcomeres produce emergent mechanical behavior in a mathematical model of musculus. PLoS Comput. Biol. Nov. 5:e1000560. doi: x.1371/journal.pcbi.1000560
PubMed Abstract | CrossRef Full Text | Google Scholar
Duvall, M. Grand., Gifford, J. 50., Amrein, Thousand., and Herzog, Due west. (2012). Altered mechanical properties of titin immunoglobulin domain 27 in the presence of calcium. Eur. Biophys. J. 42, 301–307. doi: x.1007/s00249-012-0875-8
PubMed Abstruse | CrossRef Full Text | Google Scholar
Edman, G. A. (2012). Residual forcefulness enhancement after stretch in striated muscle. A consequence of increased myofilament overlap? J. Physiol. 590(Pt half dozen), 1339–1345. doi: 10.1113/jphysiol.2011.222729
PubMed Abstract | CrossRef Full Text | Google Scholar
Edman, Yard. A. P., Elzinga, Yard., and Noble, M. I. M. (1978). Enhancement of mechanical performance past stretch during tetanic contractions of vertebrate skeletal muscle fibres. J. Physiol. 281, 139–155. doi: 10.1113/jphysiol.1978.sp012413
PubMed Abstract | CrossRef Full Text | Google Scholar
Edman, K. A. P., Elzinga, G., and Noble, M. I. Thou. (1982). Rest force enhancement after stretch of contracting frog single muscle fibers. J. Gen. Physiol. eighty, 769–784. doi: 10.1085/jgp.fourscore.five.769
PubMed Abstruse | CrossRef Full Text | Google Scholar
Epstein, M., and Herzog, Due west. (1998). Theoretical Models of Skeletal Musculus:Biological and Mathematical Considerations. New York, NY: John Wiley & Sons Ltd.
Fenn, W. O. (1923). A quantitative comparison between the free energy liberated and the work performed by the isolated sartorius of the frog. J. Physiol. 58, 175–205. doi: 10.1113/jphysiol.1923.sp002115
PubMed Abstract | CrossRef Full Text | Google Scholar
Gasser, H. S., and Colina, A. V. (1924). The dynamics of muscular contraction. Proc. R. Soc. Lond. B Biol. Sci. 96, 398–437. doi: 10.1098/rspb.1924.0035
CrossRef Total Text | Google Scholar
Gordon, A. M., Huxley, A. F., and Julian, F. J. (1966). The variation in isometric tension with sarcomere length in vertebrate musculus fibres. J. Physiol. 184, 170–192. doi: ten.1113/jphysiol.1966.sp007909
PubMed Abstract | CrossRef Full Text | Google Scholar
Herzog, Westward. (2014). Mechanisms of enhanced force production in lengthening (eccentric) musculus contractions. J. Appl. Physiol. (1985) 116, 1407–1417.
PubMed Abstract | Google Scholar
Herzog, W., Duvall, M., and Leonard, T. R. (2012a). Molecular mechanisms of muscle force regulation: a role for titin? Exerc. Sport Sci. Rev. forty, l–57. doi: 10.1097/JES.0b013e31823cd75b
PubMed Abstruse | CrossRef Total Text | Google Scholar
Herzog, W., Leonard, T., Joumaa, V., Duvall, K., and Panchangam, A. (2012b). The three filament model of skeletal muscle stability and forcefulness production. Mol. Cell. Biomech. 9, 175–191.
PubMed Abstract | Google Scholar
Herzog, W., and Leonard, T. R. (2002). Strength enhancement following stretching of skeletal muscle: a new mechanism. J. Exp. Biol. 205, 1275–1283.
PubMed Abstruse | Google Scholar
Herzog, W., and Leonard, T. R. (2006). Response to the (Morgan and Proske) Letter to the Editor by Walter Herzog and Tim Leonard (on behalf of the authors). J. Physiol 578(Pt 2), 617–620. doi: 10.1113/jphysiol.2006.125443
CrossRef Total Text
Herzog, Due west., and Leonard, T. R. (2013). Rest force enhancement: the neglected property of striated muscle contraction. J. Physiol. 591(Pt viii), 2221. doi: 10.1113/jphysiol.2012.248450
PubMed Abstract | CrossRef Full Text | Google Scholar
Huxley, A. F., and Niedergerke, R. (1954). Structural changes in musculus during contraction. Interference microscopy of living muscle fibres. Nature 173, 971–973. doi: 10.1038/173971a0
PubMed Abstract | CrossRef Full Text | Google Scholar
Huxley, H. Due east. (1953). Electron microscope studies of the organization of the filaments in striated muscle. Biochim. Biophys. Acta 12, 387–394. doi: 10.1016/0006-3002(53)90156-5
CrossRef Full Text | Google Scholar
Huxley, H. Eastward., and Hanson, J. (1954). Changes in cross-striations of musculus during contraction and stretch and their structural implications. Nature 173, 973–976. doi: 10.1038/173973a0
PubMed Abstract | CrossRef Total Text
Joumaa, 5., and Herzog, W. (2013). Energy cost of force product is reduced later active stretch in skinned musculus fibres. J. Biomech. 46, 1135–1139. doi: 10.1016/j.jbiomech.2013.01.008
PubMed Abstract | CrossRef Full Text | Google Scholar
Julian, F. J., and Morgan, D. Fifty. (1979). The effects of tension on non-uniform distribution of length changes practical to frog musculus fibres. J. Physiol. 293, 379–392. doi: x.1113/jphysiol.1979.sp012895
PubMed Abstruse | CrossRef Full Text | Google Scholar
Labeit, D., Watanabe, K., Witt, C., Fujita, H., Wu, Y., Lahmers, S., et al. (2003). Calcium-dependent molecular bound elements in the behemothic protein titin. Proc. Natl. Acad. Sci. U.S.A. 100, 13716–13721. doi: 10.1073/pnas.2235652100
PubMed Abstruse | CrossRef Full Text | Google Scholar
Lee, Due east. J., and Herzog, W. (2008). Residual force enhancement exceeds the isometric force at optimal sarcomere length for optimized stretch conditions. J. Appl. Physiol. 105, 457–462. doi: x.1152/japplphysiol.01109.2006
PubMed Abstract | CrossRef Full Text | Google Scholar
Lee, H. D., and Herzog, Due west. (2002). Force enhancement post-obit muscle stretch of electrically and voluntarily activated homo adductor pollicis. J. Physiol. 545, 321–330. doi: 10.1113/jphysiol.2002.018010
PubMed Abstruse | CrossRef Full Text | Google Scholar
Leonard, T. R., Duvall, M., and Herzog, Due west. (2010). Force enhancement post-obit stretch in a unmarried sarcomere. Am. J. Physiol. Cell. Physiol. 299, C1398–C1401. doi: x.1152/ajpcell.00222.2010
PubMed Abstract | CrossRef Full Text | Google Scholar
Leonard, T. R., and Herzog, W. (2010). Regulation of musculus strength in the absence of actin-myosin based cross-bridge interaction. Am. J. Physiol. Cell. Physiol. 299, C14–C20. doi: 10.1152/ajpcell.00049.2010
PubMed Abstract | CrossRef Full Text | Google Scholar
Llewellyn, K. Due east., Barretto, R. P. J., Delp, Southward. L., and Schnitzer, G. J. (2008). Minimally invasive high-speed imaging of sarcomere contractile dynamics in mice and humans. Nature 454, 784–788. doi: 10.1038/nature07104
PubMed Abstract | CrossRef Full Text | Google Scholar
Lundsgaard, E. (1930). Untersuchungen über Muskelkontraktionen ohne Milchsäurebildung. Biochem. Z. 217, 162–177.
Morgan, D. Fifty. (1994). An explanation for balance increased tension in striated muscle after stretch during contraction. Exp. Physiol. 79, 831–838. doi: 10.1113/expphysiol.1994.sp003811
PubMed Abstruse | CrossRef Full Text | Google Scholar
Morgan, D. 50., Mochon, Southward., and Julian, F. J. (1982). A quantitative model of intersarcomere dynamics during fixed-end contractions of single frog musculus fibres. Biophys. J. 39, 189–196. doi: x.1016/S0006-3495(82)84507-4
PubMed Abstract | CrossRef Total Text | Google Scholar
Morgan, D. Fifty., and Proske, U. (2006). Can all residual force enhancement be explained by sarcomere non-uniformities? J. Physiol. 578(Pt 2), 613–615. doi: ten.1113/jphysiol.2006.125039
PubMed Abstract | CrossRef Full Text | Google Scholar
Morgan, D. L., Whitehead, North. P., Wise, A. Thousand., Gregory, J. East., and Proske, U. (2000). Tension changes in the cat soleus muscle following slow stretch or shortening of the contracting muscle. J. Physiol. 522(Pt 3), 503–513. doi: x.1111/j.1469-7793.2000.t01-two-00503.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Oskouei, A. East., and Herzog, Due west. (2005). Observations on forcefulness enhancement in sub-maximal voluntary contractions of human adductor pollicis muscle. J. Appl. Physiol. 98, 2087–2095. doi: 10.1152/japplphysiol.01217.2004
PubMed Abstract | CrossRef Full Text | Google Scholar
Peterson, D., Rassier, D. East., and Herzog, W. (2004). Strength enhancement in single skeletal musculus fibres on the ascending limb of the force-length human relationship. J. Exp. Biol. 207, 2787–2791. doi: 10.1242/jeb.01095
PubMed Abstract | CrossRef Full Text | Google Scholar
Pollack, G. H. (1990). Muscles & Molecules - Uncovering the Principles of Biological Motion. Seattle, WA: Ebner and Sons.
Rassier, D. E., Herzog, W., and Pollack, One thousand. H. (2003a). Dynamics of individual sarcomeres during and after stretch in activated single myofibrils. Proc. Biol. Sci. 270, 1735–1740. doi: 10.1098/rspb.2003.2418
PubMed Abstract | CrossRef Full Text | Google Scholar
Rassier, D. East., Herzog, W., and Pollack, G. H. (2003b). Stretch-induced force enhancement and stability of skeletal muscle myofibrils. Adv. Exp. Med. Biol. 538, 501–515. doi: ten.1007/978-1-4419-9029-7_45
PubMed Abstruse | CrossRef Full Text | Google Scholar
Rassier, D. Eastward., Herzog, W., Wakeling, J. K., and Syme, D. (2003c). Stretch-induced, steady-state strength enhancement in unmarried skeletal muscle fibers exceeds the isometric forcefulness at optimal fibre length. J. Biomech. 36, 1309–1316.
Google Scholar
Rayment, I., Holden, H. M., Whittaker, 1000., Yohn, C. B., Lorenz, Chiliad., Holmes, G. C., et al. (1993). Construction of the actin-myosin complex and its implications for musculus contraction. Scientific discipline 261, 58–65. doi: ten.1126/scientific discipline.8316858
PubMed Abstract | CrossRef Full Text | Google Scholar
Schachar, R., Herzog, Westward., and Leonard, T. R. (2004). The furnishings of muscle stretching and shortening on isometric forces on the descending limb of the force-length human relationship. J. Biomech. 37, 917–926. doi: 10.1016/j.jbiomech.2003.10.006
PubMed Abstruse | CrossRef Full Text | Google Scholar
Schappacher-Tilp, M., Leonard, T., Desch, G., and Herzog, W., (2015). A novel iii-filament model of force generation in eccentric contraction of skeletal muscles. PLoS. 1 10:e0117634. doi: 10.1371/journal.pone.0117634
PubMed Abstract | CrossRef Full Text | Google Scholar
Sugi, H., and Tsuchiya, T. (1988). Stiffness changes during enhancement and deficit of isometric force by wearisome length changes in frog skeletal musculus fibres. J. Physiol. 407, 215–229. doi: 10.1113/jphysiol.1988.sp017411
PubMed Abstruse | CrossRef Full Text | Google Scholar
Zahalak, One thousand. I. (1997). Can musculus fibers be stable on the descending limbs of their sarcomere length-tension relations? J. Biomech. thirty, 1179–1182.
PubMed Abstract | Google Scholar

Which Part Of The Sarcomere Stays Almost The Same Size During Contraction,
Source: https://www.frontiersin.org/articles/10.3389/fphys.2015.00174/full
Posted by: pillsburyourt1996.blogspot.com
0 Response to "Which Part Of The Sarcomere Stays Almost The Same Size During Contraction"
Post a Comment